 欢迎来到乐鱼网页版登录入口网站!欢迎来到乐鱼网页版登录入口网站!
欢迎来到乐鱼网页版登录入口网站!欢迎来到乐鱼网页版登录入口网站!
15221734409



期刊:Nature Chemical Biology
年份:2025
核心定位:澄清 lactylation 异构体争议,建立“Kl-la 是组蛋白糖酵解响应乳酰化”的分析标准。
一页读懂:这篇文章解决什么问题
此前争议:
1. Kₗ-la、Kᴅ-la 与 Kce 分子量相同/相近,常规 MS 容易混淆。
2.乳酸、MGO、LGSH 均与糖酵解有关,但对应的是不同化学来源。
3. 社区质疑:组蛋白 Kla 到底是酶促 L-乳酰化,还是非酶促副产物?
作者结论:
1.细胞组蛋白上的主要乳酰化异构体是 Kl-la。
2. Kl-la 随糖酵解/葡萄糖动态升高;Kd-la 与 Kce 在野生型细胞中不构成主要组蛋白标记。
3. lactyl-CoA 与 Kl-la 水平正相关,支持其作为高能中间体。
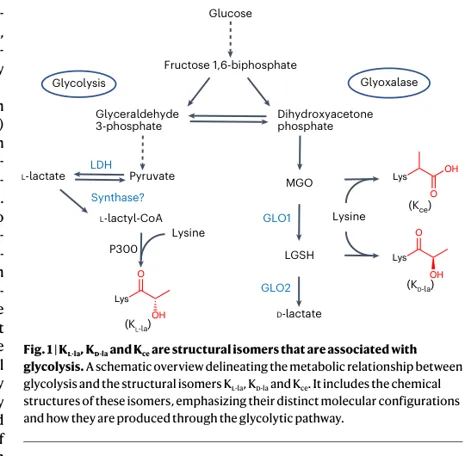
研究背景:乳酸相关修饰存在“三种异构体”
关键点:三种修饰都“连接糖酵解”,但生物学含义不同。本文的核心贡献是把“同名 Kla”拆分为可验证的化学实体。
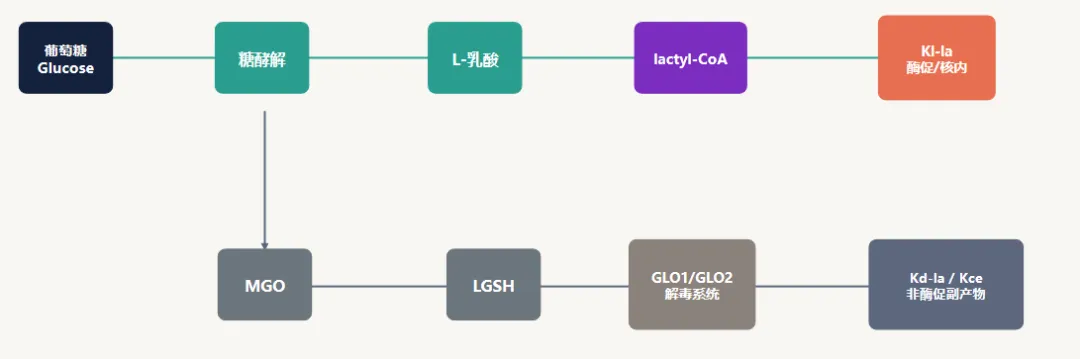
总体技术路线:用正交证据拆分异构体
解读:文章强在“方法学闭环”,不是依赖某一个抗体或某一次质谱结果,而是用免疫学、分析化学、代谢示踪和遗传/药理扰动相互验证。
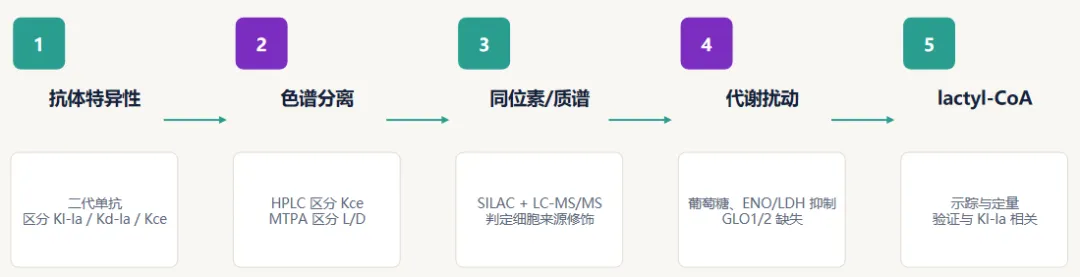
结果 1:PTM 特异性抗体可区分三种修饰
实验设计:
1.合成三类抗原:修饰肽库、修饰 BSA、序列特异性组蛋白肽。
2.分别检测 anti-Kl-la、anti-Kd-la、anti-Kce 的交叉反应。
3.每种抗体对目标 PTM 至少有约 50 倍偏好。
解读
1.这一步解决“抗体是否把三种异构体混在一起识别”的问题。
2.但抗体证据本身仍不够,必须用色谱/质谱进一步确认。
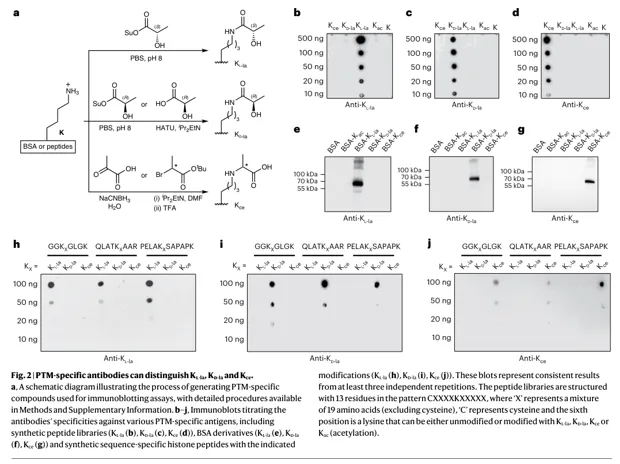
结果 2:HPLC + 手性衍生实现异构体分辨
1.Kce 与 Kl/d-la 可由反相 HPLC 区分:Kce 与 Kl-la/Kd-la 虽然质荷比相同/相近,但肽段保留时间不同。
2.Kl-la 与 Kd-la 需要 MTPA-Cl 手性衍生:L/D 仅差一个手性中心,普通 C18 难分离;衍生后差异被放大。
3.单氨基酸层面确认 L/D 归属:先氨肽酶消化到修饰氨基酸,再 LC-MS/MS 判定。
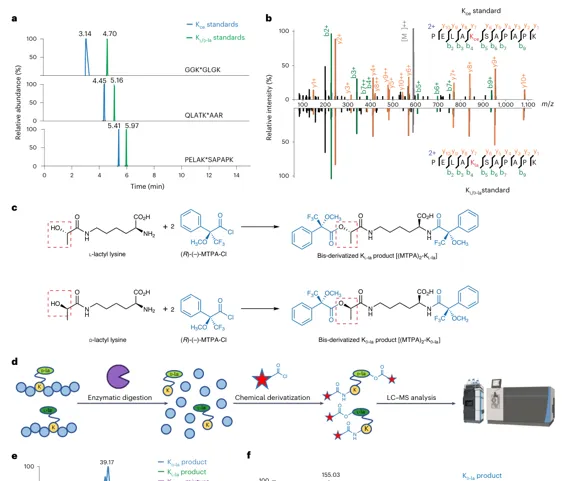
结果 3:细胞组蛋白乳酰化主要是 Kl-la
证据链:
1.MCF-7 细胞用重同位素赖氨酸标记,提取组蛋白。
2.亲和富集目标 PTM 肽段后加入轻标合成标准肽。
3.细胞来源肽与 Kl/d-la 标准共洗脱,而不与 Kce 标准共洗脱。
4.进一步经 MTPA 衍生,细胞来源修饰与 Kl-la 标准一致,而非 Kd-la。
结论:在野生型细胞组蛋白中,Kl-la 是主要乳酰化异构体;Kd-la/Kce 未被高灵敏度质谱检出。
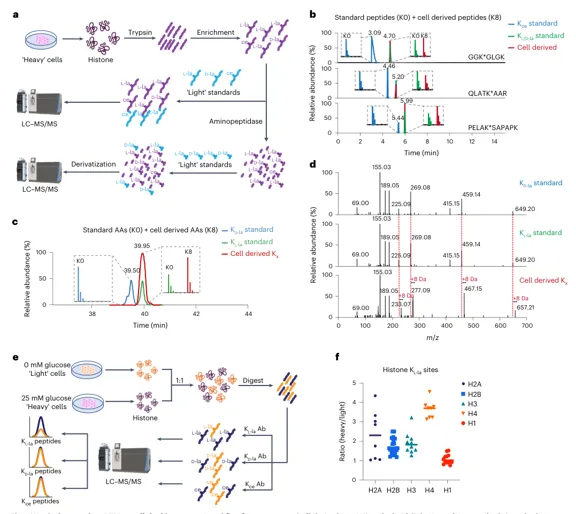
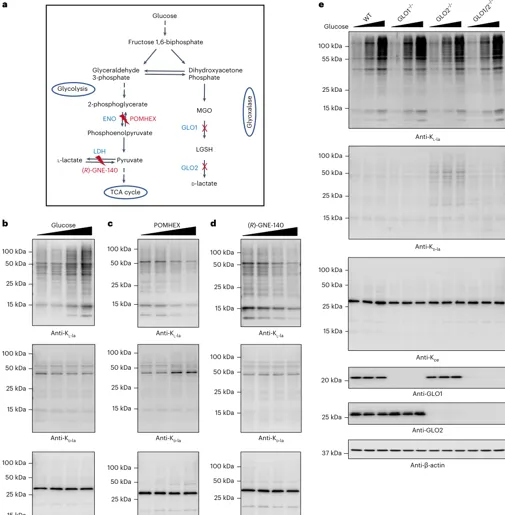
结果 4:糖酵解选择性驱动 Kl-la
关键扰动与趋势:
1.葡萄糖升高:Kl-la 剂量依赖性升高;Kd-la/Kce 基本不响应
2.ENO 抑制 POMHEX:Kl-la 下降;Kd-la 上升,提示碳流偏向 glyoxalase 相关路径
3.LDH 抑制 GNE-140:Kl-la 下降;Kd-la/Kce 基本不变
4.GLO1/2 缺失:GLO2 缺失可诱导 Kd-la;Kl-la 仍主要由糖酵解响应调控
解读:Kl-la 更像“糖酵解—乳酸—lactyl-CoA”通路输出;Kd-la/Kce 更像糖酵解副产物逃逸解毒后的化学修饰。
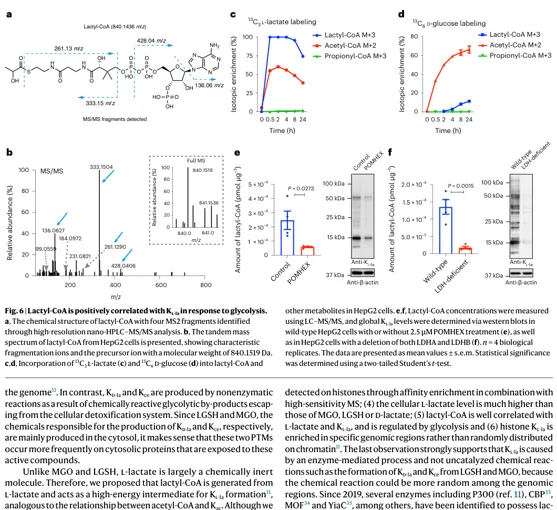
结果 5:lactyl-CoA 与 Kl-la 水平正相关
作者想回答的问题:
1.L-乳酸本身化学惰性较强,如何形成赖氨酸乳酰化?
2.是否存在类似 acetyl-CoA 的高能中间体?
3.lactyl-CoA 是否随糖酵解变化并与 Kl-la 同步?
主要证据:
1.高分辨 LC-MS/MS 在细胞中确认 lactyl-CoA。
2.13C3-L-lactate 与 U-13C6-glucose 均可标记 lactyl-CoA。
3. POMHEX、LDH 缺失或 LDH 抑制降低 lactyl-CoA,并同步降低 Kl-la。
整合模型:酶促 Kl-la 与非酶促 Kd-la/Kce 需要分开理解
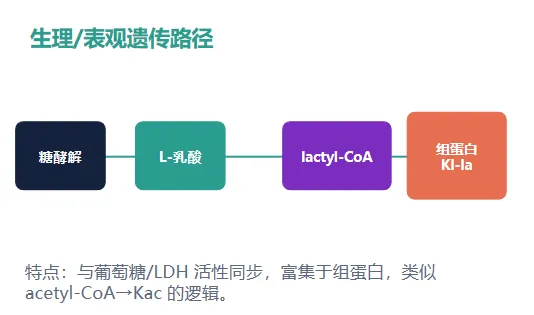
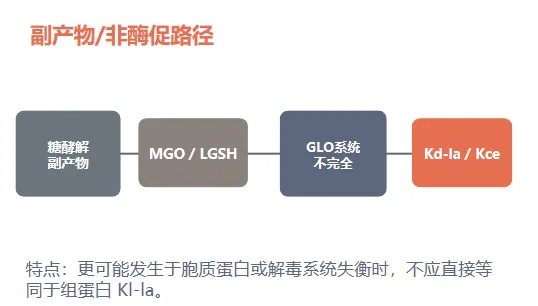
本文的概念推进:把“乳酰化”从一个泛化标签,推进到化学结构、代谢来源和细胞定位均可检验的 PTM 实体。
文章创新性:方法学校正比单一机制发现更重要
1.方法创新:建立异构体特异性抗体;用 MTPA-Cl 手性衍生解决 Kl-la/Kd-la 分辨问题。
2.概念创新:证明糖酵解动态响应的组蛋白乳酰化主要是 Kl-la,而非 Kd-la 或 Kce。
3.代谢连接:lactyl-CoA 的检测、示踪和代谢扰动支持其作为 Kl-la 形成中间体。
4.领域规范:提示未来乳酰化研究必须说明检测的是哪一种异构体,避免 pan-Kla 过度解释。
评价:这篇文章的价值在于为“乳酰化—糖酵解—表观遗传”研究提供了更严格的检测边界。
关键局限与未解问题
1.dactyl-CoA 的合成酶是谁?
文章确认其存在和代谢相关性,但未明确哺乳动物细胞中负责生成lactyl-CoA的酶。
2.KI-la 的reader是否特异?
已有 writer/eraser 线索,但不同KI-la位点的识别蛋白和转录输出仍需系统解析。
3.低丰度Kd-la/Kce是否完全没有功能?
作者未检测到主要组蛋白Kd-la/Kce,但不能排除极低丰度、位点特异性调控。
4.非组蛋白KI-la的功能深度不足
本文重点是异构体鉴定和糖酵解响应,不是逐一解析靶蛋白功能。
对后续课题的启示:乳酰化研究应如何做得更稳
1.筛选:pan-Kla WB/IF修饰组学初筛
2.确认:IP-WB + LC-MS/MS明确 Kl-la 位点
3.异构体:标准肽/手性衍生排除 Kd-la/Kce 混淆
4.因果:葡萄糖/乳酸/LDHPOMHEX/GNE-140
5.功能:K→R/K→Q 突变表型救援
适用场景:肿瘤耐药、巨噬细胞/Treg 免疫代谢、纤维化、缺氧微环境、代谢性疾病中的“乳酸—表观遗传—表型”机制。
Take-home messages:
1.Kl-la 是组蛋白糖酵解响应乳酰化的主要形式。
2.Kd-la 与 Kce 更偏向糖酵解副产物逃逸解毒后的非酶促修饰。
3.仅用 pan-Kla 抗体不足以支持严谨机制结论。
4.高水平乳酰化研究需要“异构体确认 + 位点质谱 + 功能突变救援”。
一句话评价:这篇文章把乳酰化研究从“是否升高”推进到“是哪一种化学修饰、由哪条代谢路径驱动、是否具有可验证因果关系”。

